4.1 碳水化物的分类、结构和性质
4.1.1 碳水化物的分类
碳水化物分单糖、双糖、低聚糖、多糖四类。糖的结合物有糖脂、糖蛋白、蛋白多糖三类。
(1)单糖不能水 解成更简单的糖的碳水化物叫单糖。单糖有3到7个碳原子,依碳原子数的多少,依次称为丙、丁、戊、已、庚糖。丙糖和丁糖以中间代谢物的形式存在,自然界存在最多是戊糖和已糖。
单糖具有醛基或酮基。有醛基者称醛糖,有酮基者称酮糖。
(2)双糖每分子能水解成两分子单糖的碳水化称双糖。营养上有意义的双糖有蔗糖、麦芽糖、乳糖三种。
(3)低聚糖每分子水解成3~8个分子单糖的碳水化物称低聚糖,也有人把水解成3~10个,甚至20个分子单糖的碳水化物归入这一类。可再分两类:水解产生的所有糖分子都是葡萄糖的称麦芽糖低聚糖,由3个葡萄糖分子组成的叫麦芽三糖,四个葡萄糖分子组成的叫麦芽四糖等等。另一类水解时产生不止一种单糖,称杂低聚糖。如大豆中的杂低聚糖水解产生棉子糖和木苏糖等,人不易消化,无法利用。但机体自己合成的杂低聚糖,有很重要的生理功用。
(4)多糖每分子更多分子的单糖的碳水化物是多糖。可再分α葡聚糖、β-葡聚糖、杂多聚糖三类。α-和β-葡萄聚糖都是葡萄糖分子组成的,但联结的方式不同,前者是以α-糖苷键联结的,后者则用β-糖苷键。α-葡聚糖有淀粉、糊精、糖原三类。组成杂多聚糖的不止一种单糖,膳食纤维属这一类,除纤维素外,膳食纤维还包括半纤维素、果胶、藻类、木质素。
(5)糖脂糖和脂质的结合物称糖脂。简单可分两类:脑苷脂和神经节苷脂。
(6)糖蛋白和蛋白多糖广义讲,碳水化物和蛋白质结合后的化合物都是糖蛋白。糖蛋白的种类很多,并有广泛的生理功用。30年代以来,好几个科学家提出了不同的分类方法。
1946年Stacey把蛋白质含量低,以碳水化物的化学反应为主的称粘多糖;蛋白含量高,化学反应以蛋白质为主的称粘蛋白。
1962年Gottschalk根据碳水化合的结构分成两大类。简言之,有重复出现的双糖单位的低聚糖链或多糖链,与蛋白质结合者称多糖蛋白,后人把它叫做Protegolycans,这个名字一直沿用至今,译作蛋白多糖。以没有重复出现双糖单位的低聚糖单位的低聚糖链与蛋白质相结合的称糖蛋白。
80年代以来的分类,从化学结构出发,分得很细,不予介绍了。本章的讨论,以Gottschalk的分类法为主。
粘多糖和粘蛋白的名字,现在营养学书上仍有出现,但含义与Stacey提出的略有不同。粘多糖(Mucopolyscaccharides)指Gottschalk分类的蛋白多糖中的碳水化物部分,或称氨基多糖(Glycosaminoglycans)。粘蛋白指Gotrtschalk分类中的糖蛋白。有时把糖脂、糖蛋白和蛋白多糖一起,统称糖结合物(Glycoconjugates)。
4.1.2 碳水化物的化学组成和结构
(1)单糖单糖中的最重要的是葡萄糖,它的结构式有五种表示方法,见图4-1。
葡萄糖有六个碳原子,以阿拉伯数字计数如图。在图4-1(A)中可看出第2、3、4、5个碳原子所连的四个基都不相同,是不对称碳原子。每个不对称碳原子有两个异构体,羟基在左面或右面代表两个不同的化合物。习惯上把与伯醇基[图4-1(A)中第6个碳原子]相连的碳原子的羟基在右面的称D系的糖。图4-1(A)中用虚线框出的部分是D系糖必备的结构。所以,图4-1(A)代表的是D-葡萄糖。
图4-1(A)所示的开链式结构可以环合起来,即第5个原子上羟基的氢移到第1个碳原子的羟基氧上,然后连成环,见图4-1(B)。这样,第1个碳原子也变成不对称的的。羟基在右面的称α,D-葡萄糖,羟基在左面的称β,D-葡萄糖。
但是六个碳原子是不在一条直线上的,六个碳原子的环是六角形的。为了区别不对称碳原子的构型,可把这个环表示如图4-1(C)。环的平面垂直纸面,粗线表示在环平面前面的链,细线代表的链在环平面的后面。图4-1(B)中右侧的
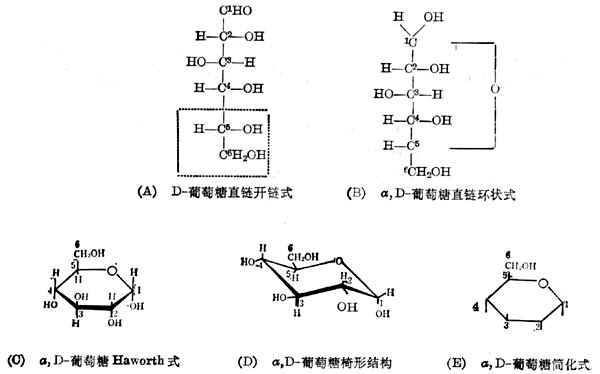
图4-1 葡萄糖的结构式
羟基写在环的平面之下,左侧的羟基写在环的平面之上。这样表示的结构式叫做Haworth投影式。
事实上,环的六个碳原子并不在一个平面上,X线衍射的结果说明它们是以椅形存在的,如图4-1(D)。但这样的表示方法书写困难,所以一般采用Haworth投影式。并且常简化如图4-1(E),这里把碳和氢都省了,仅用一条垂线表示羟基。现在国际上有一种通用的符号来代表糖及其衍生物,葡萄糖的是Glc。
自然界存在的葡萄糖,不论在单糖、双糖、低聚糖或多糖中,都以图4-1(D)的形式存在。只有在水溶液中,才有约1%以开链的形式存在。
糖有旋光性。在旋光计中,有的能使偏振光的平面右旋,有的使之左旋。命名时,右旋用(+)表示,左旋用(-)表示。不是所有的D系的糖都是右旋的。α,D-葡萄糖是右旋的,它的全名是α,D(+)葡萄糖。一般提到的葡萄糖,除特别指出的以外,都是α,D(+)葡萄糖。
果糖是D系的、β异构体、左旋的,全名是β,D(-)果糖。它的环由五个碳原子组成,在同一平面上。
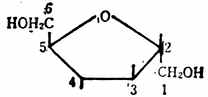
图4-2 β,D(-)果糖
半乳糖是葡萄糖的异构体,所不同的是第4个碳原子上的羟基在环的上面,而葡萄糖的在环的下面。符号Gal。
(2)双糖在葡萄糖的环状结构中,第一个碳原子上有一个羟基。它可和另一个有羟基的化合物失水而成为糖苷,形成的键叫糖苷键。
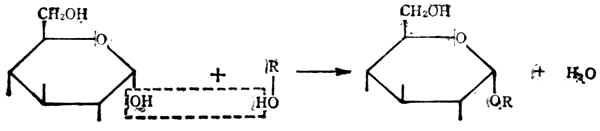
葡萄糖 葡萄糖苷
糖苷是自然界存在的一大类化合物,有些可作为药物。
如ROH是一个单糖分子,这样结合的就成为双糖。如两个单糖分子都是葡萄糖,以α-1,4糖甘键(即第一个葡萄糖分子的第1个碳原子上的羟基和第二个葡萄分子的第4个碳原子上的羟基脱水相结合而形成的键,亦写作α1→4)结合后得到的是麦芽糖。
蔗糖是α,D-葡萄糖和β,D-果糖以α-1→2糖苷键结合的。乳糖是β,D-半乳糖以β-1→4糖苷键和α,D-葡萄糖相结合的。
(3)低聚糖 麦芽三糖到麦芽八糖,都是α,D-葡萄糖以α-1→4和α-1→6糖苷键结合的。杂低聚糖匠结构比较复杂,将于糖蛋白中加以介绍。
(4)多糖
①淀粉淀粉是由许多葡萄糖分子组成的。与水调成糊状并加热到60~80℃后,可分成直链淀粉和支链淀粉两部分。前者约占15%,后者约占85%。
直链淀粉也由葡萄糖分子,以α-1→4苷键连成一条直链,并蜷曲如图4-3。
支链淀粉也由葡萄糖分子组成,它们不仅以α-1→4糖苷键连成直链,还有分支。分支处是以α-1→6糖苷键结合的。每一支有20~30个葡萄糖分子。图4-4是它的示意图。图4-5是图4-4中框出部分的放大,用以表示葡萄分子间的联结方式。
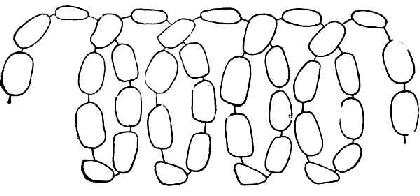
图4-3 直链淀粉示意图
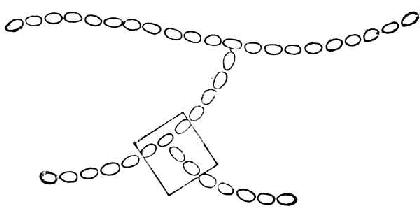
图4-4 支链淀粉示意图
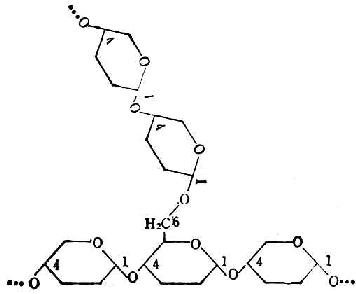
图4-5 葡萄糖分子在支链淀粉中的结合方式
②糖原糖原由6000~3000个葡萄糖分子组成,结构式和支链淀粉相似,不过分支更多。每一分支约有11~18个葡萄糖分子。分支处是α1→6糖苷键,其余是α1→4糖苷键。
③糊精和麦芽低聚糖二者都是淀粉水解的中间产物。结构中有α1→4和α1→6糖苷键。
④纤维素纤维素是β-葡萄糖以β1→4糖苷键连成直链,每链约有2800个葡萄糖分子。结构如图4-6,图中葡萄糖分子的环,是前、后交替地与纸的平面垂直的。
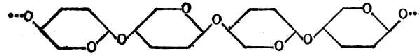
图4-6 纤维素的结构式
⑤膳食纤维
半纤维素是D-甘露糖、D-木糖、D-葡萄糖、D-半乳糖以β1→4糖苷糖连起来的,分支上有阿拉伯糖、半乳糖和葡萄糖醛酸。
果胶主链葡萄糖醛酸连成的,侧链上有鼠李糖、阿拉伯糖、岩藻糖、木糖。
藻类多糖主链由甘露糖、木糖、葡萄糖醛酸和葡萄糖组成,侧链有半乳糖。
木质素木质不是碳水化物,是苯丙烷的聚合物。由于和其余膳食纤维同时存在,且有类似的功用,所以把它作为膳食纤维中的一类。
膳食纤维和粗纤维不同。植物性食物中,有胃肠道不能消化的物质,统称膳食纤维;食物用酸碱处理后的不溶物称为粗纤维。经过这一处理,大部分膳食纤维都丢失,测得的数值仅有膳食纤维总量的20~50%。
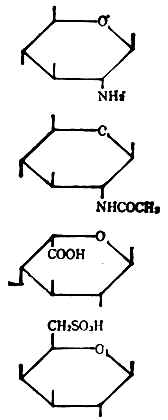
⑥单糖的衍生物低聚糖和多糖不仅能由单糖组成,还能由单糖的衍生物所组成。表4-1列出了它们的类别,每类举一例表示结构式及其名称和符号。
表4-1 单糖的衍生物
| 类别 | 结构式 | 名称 | 符号 |
| 氨基糖 |
| β-D氨基葡萄糖 | ΒDGlen |
| 乙酰氨基糖 | N-乙酰氨基-β-D葡萄 | ΒDGlcNAc | |
| 糖醛酸 | α-L-艾杜糖醛酸 | Αltdu | |
| 硫酸酯 | 4-硫酸-β-D-半乳糖 | ΒDGal-6SO3h |
另有一类组成低聚糖和多糖的单糖衍生物,叫做唾液酸,除有乙酰氨基和羟基外,还和甘油连在一起。结构式见图4-7,符号NeuAc。
糖的醛基被氧化后,得糖酸如D-葡萄酸(图4-8)。醛基被还原成羟基后,得糖醇如山梨醇(图4-9)。
⑦糖脂糖脂的基本结构是鞘氨醇(图4-10),它的氨基与脂肪酸(含14~16个碳原子)酰化后得脂酰鞘氨醇,符号为Cer。再与葡萄糖或半乳糖相连成脑苷脂。和低聚糖(由几个或多至30多个单糖或其衍生物组成)相连后成神经节苷脂。已确定结构式的有数十种,区别在于糖链和脂肪酸的不同。
 | 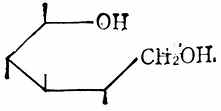 | 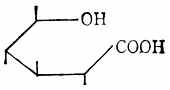 |
| 图4-7 唾液酸 | 图4-8 D-葡萄酸 | 图4-9 山梨醇 |
糖脂的糖链,和蛋白多糖的糖链常有一部分相同。所以它们的生理功用,有不少相似之处。
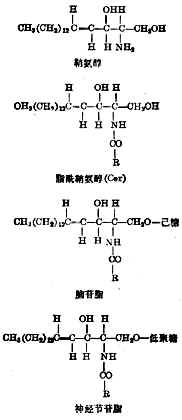
图4-10糖脂的化学结构
⑧糖蛋白中低聚糖糖蛋白中的单糖或基衍生物有以下种:
戊糖:阿拉伯糖(Ara)、木糖(Xy1);
甲基戊糖:岩藻糖(Fuc);
已糖:葡萄糖、半乳糖、甘露糖(Man);
乙酰氨基已糖:乙酰氨基葡糖(GlcNAc)、乙酰氨基半乳糖(GallNAc);
唾液酸。
不同的糖蛋白,由这些单糖或其衍生物中的一部分,组成不超过15个单体的低聚糖链,再和蛋白质结合而成。糖含量为1~85%。
⑨蛋白多糖中的聚氨基糖这一类糖是多糖。糖链中有交替出现的糖醛酸(葡萄醛酸或艾杜糖醛酸)和氨基已糖(氨基葡萄糖或氨基半乳糖或基乙酰化产物)。习惯上说成有交替出现的双糖单位。共分七类,见表4-2。
在这七类中,透明质酸有羧基,其它六类都有硫酸基,因此聚氨基糖都是多价的阴离子。
究竟有多少单糖分子聚合成聚氨基糖?根据来源而不同,分子量自4000到1000000以上。
表4-2 蛋白多糖中的氨基糖的分类和组成
| 聚氨基糖 | 双糖单位 | 取代基 | 其它单糖 | |
| 糖醛酸 | 氨基已糖 | |||
| 透明质酸 | βDGel | βDGleNAc | Gal;Xyl | |
| 4-硫酸软骨素 | ΒDGel | ΒDGalNAc | GalNAc 6位硫酸化 | Gal;Xyl |
| 6-硫酸软骨素 | ΒDGel | βDGalNAc | GalNAc 6位硫酸化 | |
| 硫酸皮肤素 | βDGal αLIeU | αDGlcN | 部分IdU2位硫酸化 部分GalNAc4或6位硫酸化; | Gal;Xyl |
| 硫酸类肝素 | ΒDGal αLIdU | αDGlcN | GlcN氨基乙酰化或硫酸化; 部分GlcN6位硫酸化; 部分IdU2位硫酸化 | |
| 肝素 | βDGal `αLIdU | αDGlcN | GlcN氨基乙酰化或硫酸化; 部分GlcN3或6位硫酸化; 部分IdU2位硫酸化 | Gal;Xyl |
| 硫酸角质素 | βDGal | βDGlcNAc | 部分GleNAc6 | GalN;Man; Fuc;NeuAc |
透明质酸单独存在,一般都是单独的一条多糖链。其它六类中,除硫酸角质素外,都通过半乳糖-木糖和蛋白质的丝氨酸相连结的,一条肽链上可连几条糖链。
4.1.3 碳水化物的物理性质
(1)溶解度单糖、双糖、低聚糖、糊精都溶于水。淀粉不溶,与水加热后可吸水膨胀,变成糊状。淀粉经酸处理生成可溶淀粉。糖原能分散在水中得乳白色胶态“溶液”。纤维素不溶。果胶能溶于水。其余膳食纤维可吸水膨胀,吸水量依来源、周围液体的PH和离子浓度等而不同。如麦麸可吸收5倍生本身重量的水。吸水后膳食纤维呈海绵状,细菌和一些分子能穿插进去。糖脂糖的一端亲水,脂质的一端疏水。糖蛋白和蛋白多糖的糖也都亲水。
(2)甜度如以蔗糖的甜度作100,一些糖及糖醇的甜度见表4-3。其中转化糖是蔗糖的水解产物,含葡萄糖和果糖各50%。
表4-3 糖的甜度
| 糖或其衍生物 | 甜度 |
| 果糖 | 173 |
| 转化糖 | 130 |
| 蔗糖 | 100 |
| 葡萄糖 | 74 |
| 山梨醇 | 60 |
| 甘露醇 | 50 |
| 半乳糖 | 32 |
| 麦芽糖 | 32 |
| 乳糖 | 16 |
4.2 碳水化物的生化代谢
4.2.1 消化吸收
碳水化物要消化或单糖才吸收。消化的过程就是水解的过程。麦芽糖、乳糖、蔗糖、麦芽低聚糖都能消化。人能消化的多糖仅淀粉一种,糖原在制成食品时已不存在了。
消化从口腔开始,口腔里有唾液淀粉酶能水解交替α1→4糖甘键,但不能水解α1→6糖苷键和相邻的α1→6糖苷键。消化产物是糊精、麦芽低聚糖和麦芽糖。
胃里没有消化淀粉的酶。唾液淀粉酶的最适PH是6.6~6.8,在食糜没有被胃酸中和以前,能持续作用一段时间,使淀粉和低聚糖能再消化一部分。
小肠内有胰液的α-淀粉酶,其作用和唾液淀粉酶相同,把直链淀粉消化成麦芽糖和麦芽三糖,支链淀粉消化成麦芽糖、麦芽三糖及由4~9个葡萄糖分子组成的而有α1→6苷键的麦芽低聚糖。
肠粘膜上皮细胞中有吸收细胞,每一细胞约有3000条微绒毛,微绒毛间的空间的有效半径约0.4nm。只有上述消化产物能够通过,与微绒毛膜上的酶反应。膜上的酶有四种:①α1→4糖苷酶,把葡萄糖分子自上述产物一个个地切下来;②异麦芽糖酶,水解麦芽低聚糖的α1→6糖苷键;③蔗糖酶,消化蔗糖;④β-半糖苷酶,消化乳糖。
所以消化分两步进行:①肠腔内的消化,产物是双糖和麦芽低聚糖;②微绒毛膜上的消化,产物是单糖。
微绒毛双脂质层的(详后)。四种酶嵌在双脂质层内,活性位伸在膜外。在其近处,还有全部嵌在膜内的运输单糖的蛋白质,这样,消化的最终产物立刻可以被运输蛋白所结合。
运输蛋白在结合葡萄糖以前,先结合肠腔内的Na+排入肠腔,肠腔中Na+都带入细胞内,释放到胞浆中。Na+排入肠腔,肠腔中的Na+浓度比细胞内高,自低浓度排到高浓度要消耗能量,所需能量由ATP供应。糖进入细胞后,约有15%流回肠腔,25%扩散入血,60%与靠近基膜一端的质膜上的另一载体蛋结合而离开细胞。这一结合不需Na+,而且运输葡萄糖的速度比葡萄糖从肠腔进入吸收细胞的速度快,所以葡萄糖不会在吸收细胞中蓄积,从而提高了吸收效率。当食糜到达空肠下部时,95%的碳水化物都被吸收了。
4.2.2 中间代谢概述
小肠吸收的碳水化物主要是葡萄糖、果糖、半乳糖三种,经门静脉送到肝脏。葡萄糖进入肝细胞后与磷酸反应生成葡萄糖-6-磷酸,这样细胞内的葡萄糖浓度可维持在低水平,使葡萄糖不断进入肝细胞。在吸收的葡萄糖中,60%以上在肝内代谢,其余入大循环。果糖和半乳糖在肝中转变为葡萄糖。
葡萄糖在肝内经分解代谢提供机体所需要的能量,多余的合成糖原保留在肝内,再有多余就转变成脂肪运送到脂肪组织储存起来。
葡萄糖的分解代谢分三个阶段:
(1)酵解葡萄糖分解的第一阶段是生成丙酮酸。这一系列反应和碳水化物在体外经酵母的作用生成丙酮酸相同,所以称为酵解。所不同的是发酵时丙酮酸进一步生成洒精;在体内,有氧时丙酮酸氧化成二氧化碳和水,无氧时还原成乳酸。
酵解分十步进行(图4-11)。为了要把葡萄糖的6个碳原子一分为二,先使它两端磷酸化,才能断列成两个接近相等的部分,然后脱去磷酸成丙酮酸。
上述反应都在细胞浆中进行。每一步都有特定的酶参加。第①、③两步消耗2分子ATP,说明葡萄糖分解代谢开始时要用能量来推动。⑥、⑨两步各产生2分子ATP。从葡萄到丙酮酸,净生2分子ATP,酵解产生的能量不多。
细胞浆中NAD+和NADH的总量基本恒定。如所有的NAD+都变成NADH,酵解就得停止。要使葡萄继续不断氧化成酮酸,必需把NADH的氢转移到另外的受体上去。有氧时氢经呼吸链氧化成水。无氧时氢交给丙酮酸使成乳酸⑩。乳酸过多时离开细胞入血。所以严重缺氧者可发生代谢性酸中毒。
(2)丙酮氧化成乙酰酶a 有氧时,丙酮酸进入线粒体。在丙酮酸脱氢酶系的帮助下,氧化脱羟成乙酰辅酸脱氢酶系的帮助下,氧化脱羟成乙酰辅酶A,同时产生1molATP和1molNADH。维生素B1B2、烟酰胺、泛酸参加。反应是不可逆的。
(3)三羧酸循环乙酰辅酶A上的两个来自葡萄糖的碳原子,经三羟酸循环生成2molCO2,完成葡萄糖的氧化。反应在线粒体中进行。这一过程分9步(图4-12)。
三羧酸循环中产生3molNADH,1molFADH2,1molGTP。
NADH把氢通过FAD和细胞色素交给氧,同时放出能量。每molNADH通过这一系列反应所放出的能量是220kJ(52.6kCal)。这些能量以ATP的形式贮存起来。每molATP能产生30.5Kj(7.3kCal),理论上每molNADHRNUD氧化产生的能量可用来合成7molATP。这是指在标准状况(25℃,1个大气压,作用物和产物浓度都是1M/LPH7)下说的。在模拟体内的条件时,实验证明只能合成3molATP。如用反应式表示:
NADH+H++1/2O2

3ADP+3H3PO4+能量
前面一个是氧化反应,后面一个是磷酸化,二者偶合在一起,称为氧化磷酸化。
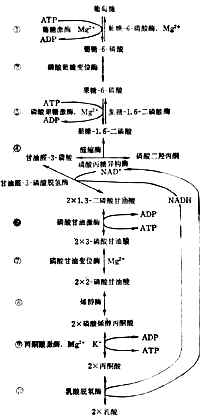
图4-11 糖的酵解
实验也证明FADH2氧化时产生2个ATP,GTP可生成1molATP。根据这些数值,葡萄糖完全氧化时,机体获得的能量可归纳如下:
①酵解:葡萄糖→2丙酮酸+8ATp
②丙酮酸氧化成乙酰辅酶A:
2丙酮酸→2乙酰辅酶A+2CO2+6ATp
③三羧酸循环:
2乙酰辅酶A→4CO2+24ATp
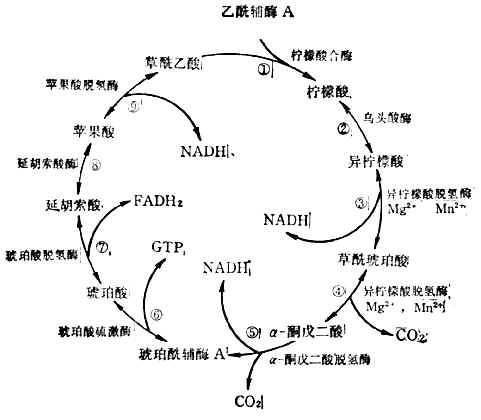
图4-12三羧酸循环
即每mol葡萄糖在体内氧化成6molCO2时可产生38molATP,机体可以利用的能量是38×30或1140kJ[(38×7.3)或277kCal]。每mol葡萄糖氧化成6molCO2和水时,自由能的改变是2870KJ(686kCal)。所以,机械效率=1140/2870×10=40%。
这是利用标准状况的数字计算的,在体内高一些,约60%。
从上面归纳的数字也可看出碳水化物提供的能量,在氧化的三个阶段中,以三羧酸循环最多。脂肪和蛋白质最后也通过三羧酸循环氧化。食物向机体提供的能量,90%来自三羧酸循环。
葡萄糖代谢另外还有一种途径,不通过果糖二磷酸,叫做已糖一磷酸通路。由于产生核榶,又叫做戊糖-磷酸通路。
(4)已糖-磷酸通路这一通路目的是提供合成核酸所需的核糖,以及合成脂肪酸、胆固醇等所需的能量供应者NADPH(还原型辅酶Ⅱ)(图4-13)。肝脏中约有30%的葡萄糖经过这一通过代谢。红细胞中这一通路也很活跃。它的失调可导致溶血性贫血。反应都在细胞浆中进行。
核糖-5-磷酸既可用于核酸的生物合成,又可与5-磷酸木酮糖反应生成甘油醛3-磷酸和果糖-6-磷酸。究竟走哪一条路,要看细胞本身对NADPH和核糖的需要量而定。许多细胞NADPH要得多,过剩的戊糖可经甘油醛-3-磷酸和果糖-6-磷酸酵解,或由果糖-6-磷酸转变成葡萄糖-6-磷酸而重走这一通路。
这一通路需维生素B1参加。要使此路畅通,膳食中需有足量的维生素B1。5-磷酸核酮糖转变为葡萄糖-6-磷酸的反应,可用于评价维生素B1参加。要使此路畅通,膳食中需有足量的维生素B1的营养状况。
(5)糖原葡萄糖到各个细胞发挥作用以后,多余的可储存起来。避免细胞内渗透压的升高,细胞把葡萄糖合成大分子的糖原。需要能量时糖原再分解葡萄糖。如肝糖原产生的葡萄糖,用以维持血糖的恒定。肌糖原产生的葡萄糖,直接用于供给能量。
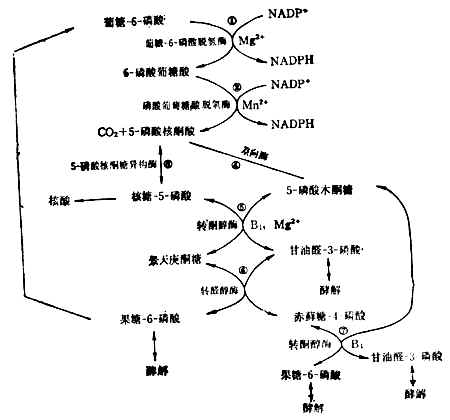
图4-13 已糖-磷酸通路
4.2.3 三大营养代谢的关系
人习惯于三餐,饱腹和空腹交替产生。所以要具备能处理碳水货物增多和缺少时保证能量供应的两套本领。
餐后血糖升沿高,胰岛素分泌增加,胰高血糖素分泌减少。使更多的葡萄糖进入肝脏、肌肉和脂肪组织;增强酶活力,加速葡萄的氧化和肝糖原、肌糖原的合成。超过糖原的储存量后,肝脏可把葡萄糖经磷酸二羟丙酮还原成甘油-3-磷酸,与乙酰辅酶A合成的脂肪酸,利用NADPH提供的能量,生成脂肪。再和蛋白质结合成极低密度脂蛋白入血,运送到脂肪组织储存。过多的碳水化物也能合成某些非必需氨基酸。
餐后约4h,不能利用食物中的葡萄糖。胰高血糖素分泌增加,胰岛素减少,转为利用肝糖原以供给血糖。肝糖原储存量不多,成人能动用的不到70g,不够一个晚上的消耗。所以,从餐后8h开始,还得利用脂肪以提供能量。甘油可变成糖,但在脂肪中所占重量不多。脂肪酸只能供能,不能转变为葡萄糖。因此,血糖的来源不得不转向氨基酸了。一般说来,除亮氨酸和赖氨酸外,其他氨基酸都是能成糖的,但从能量的观点看,只有丙氨酸和谷氨酸可作葡萄糖的来源。此时血糖的90%来自丙氨酸,10%来自谷氨酸。
肝脏能把肌肉中送来的丙氨酸作用成丙酮酸而生成葡萄糖。空腹时,肌肉中进入血液的氨基酸增加,其总量的30~40%是丙氨酸。但肌肉蛋白质的丙氨酸含量不到10%。另外的丙氨酸从哪里来的?葡萄糖经酵解可生丙酮酸,再经转氨作用丙氨酸。但这样葡萄糖的量并没有增加。增加的丙氨酸实际来自其他氨其酸。
肾脏把谷氨酸去氨后得α酮戊二酸,再和运来的成糖氨基酸如丙酸起转氨作用而得丙酮酸,最后得葡萄糖。α-酮戊二酸也能经三羟酸循环形成苹果酸,最后成葡萄糖。
| 上一页:3.5 脂类的供给量 |
| 当前页:4.1 碳水化物的分类、结构和性质 |
| 下一页:4.3 碳水化物的生理功用 |