幽门螺杆菌及其感染
1 概述
胃细菌学的研究,长期来是一个被忽视的领域。1983年Marshall和Warren从慢性活动性胃炎患者胃粘膜活检标本中分离到幽门螺杆菌(Helicobacter pylori,Hp)是对这一领域重要的突破。此后不久即在国际消化病学界引起了巨大轰动,它的发现对消化病学、特别是胃十二指肠病学的发展起了极大的推动作用。现在已经清楚它是许多慢性胃病(慢性胃炎、消化性溃疡、胃癌等)发生发展中一个重要致病因子。
1.1 Hp的历史、发现和命名
1.1.1 历史背景
Marshall在分离到Hp后又回过来探索了历史上的文献,发现这一类细菌实际上早就已经被人注意过。1893年在狗中,1896年在大鼠和猫中已有人报告在其胃中偶然见到螺形菌。本世纪初在溃疡性胃癌病人胃内容物中曾找到同样的细菌。其他报告也证实了这一些发现。也注意到健康人中未发现这些细菌。经过30年的努力,在散载的报告中,良性消化性溃疡病人胃中发现了这些细菌。1938年Doenges在一份综合性尸解研究报告中提出胃中螺形菌的流行率达43%,但是并未检查出这一细菌与不同的胃部疾病之间的关系。
关于这些细菌在人胃疾病中可能的作用一直存在着争论。有些研究者提出人们在活检标本中所见到的这种细菌是经口吞服的污染物。这一假说1959年由于当时的一位有影响的学者palmer发表了1000例胃活检标本大量组织学研究的报告而占了优势。在此以后对胃的细菌学的兴趣被泼了一盆冷水。
1975年Steer和Colin-Jones报告了胃溃疡病人中胃粘液层下的胃粘膜上发现了细菌,重新引起了人们对胃细菌在消化性溃疡致病机制中作用的兴趣,这提示细菌可能降低胃粘膜的抵抗力,因此易感于溃疡。Steer等企图分离这一细菌。结果生长的是绿脓杆菌。后来仔细审阅文章中的图片提示在粘膜中所见的是一种与绿脓杆菌无关的螺形菌。现在看来这些作者分离到的绿脓性杆菌可能是来自内窥镜的污染菌,而图片中所见的才是我们现在大家感兴趣的Hp。
现在已知Hp能产生大量尿素酶,在发现Hp之前许多学者曾证明在许多种动物胃中有内源性尿素酶活性的存在。1924年Luck和Seth最早描述了这一活性。1955年Kornber和Devies在一篇综述中下结论认为胃尿素酶主要存在于胃体部,且来源于细菌。后来在人粪中的研究还证实了这些酶与溃疡病之间的关系,甚至有人用尿素治疗某些病人取得了疗效。
随着Hp的发现,人们现在可以把这些研究放在一起得到一个清晰的认识,把胃中的Hp与尿素酶联系起来了。在这整个过程中从胃粘膜活检标本中分离Hp是一个关键因素。没有这一发现,人们对胃病的认识仍将受到限制。
1.1.2 发现和命名
在上述历史背景以及胃镜检查已大大普及、分离培养空肠弯曲菌的微需O2方法已经成熟的条件下,Marshall等敏感地意识到胃中所见的螺形菌,形态上与粪便中分离到的空肠弯曲菌相似。既然用常规的需O2和厌O2方法都不能把它分离出来,何不用分离培养空肠弯曲菌的方法试一试呢?结果因此使他们获得了成功。国内是在1985年取得成功的。最初信凭它形态上与弯曲菌属细菌相似,分离培养方法也相似,只是从胃标本中分离到的,因此称作胃弯曲菌样细菌(gastric campylobacter like organism,GCLO)。不久,由于此菌在胃窦部多见,又改称作幽门弯曲菌(campylobacter pyloridis,CP),后来又因为这一名称不符合细菌统一的拉丁文命名原则,又改为cacpylobater pylori,简写与中文译名未变。随着对这一细菌的生物学性状研究的不断深入,发现这一细菌虽在形态和培养特性上与弯曲菌属细菌有相似之处,但也有许多特性与弯曲菌属细菌明显不同。曾有人根据Hp的16SrRNA的核苷酸序列与弯曲菌属细菌不同,而与wolinella succinogenes相近,提出应归入wolinella属。1989年Goodwin等把CP与弯曲菌属与wolinella属的代表菌种从超微结构、脂肪酸组成、呼吸醌、生长特点、酶的活性等五种表现型特征上作了系统的比较。他们认为CP既不应属于弯曲菌属,亦不归Wollinella属,应另立一个新属,称作螺杆菌属(helicobacter,H)。因之,CP应相应改称为幽门螺杆菌(Helicobacter pylori, Hp)。而当时已从雪貂胃中分离到的类似Hp的一种campylobacter mustelae相应改为雪貂螺杆菌(helicobactermustelae,Hm)。自从新立螺杆菌属以来,国际上已先后有人从多种动物胃肠道分离到新的菌种认为是属于螺杆菌属的,亦有人从过去已归属于弯曲菌等属的细菌中拉出某些细菌来认为应归于螺杆菌。从现有报道看,螺杆菌属的细菌已不下10余种。其中,除了Hp和Hm以外,还有一种猫螺杆菌(helicobacter felis,Hf)亦已得到较多人的认同以外,其他的尚均还在被认同之中。
1.2 Hp感染与上消化道疾病的关系
Hp感染与上消化道疾病之间的高度相关性已有大量报道,且已取得了基本一致的认识如下。
1.2.1 Hp感染与慢性胃炎
Hp与慢性浅表性胃炎之间的病因关系已充分确立。其证据如下:①事实上所有Hp阳性者都证实有胃窦炎。②若Hp感染经抗菌治疗,Hp根除后,胃炎可消退。③在一些动物模型中,接种由患者胃中分离而得到的Hp后,可复制出慢性浅表性胃炎的病损。④Marshall和Morris,分别用患者的Hp对自己作了自身感染,亦取得了同样结果。
1.2.2 Hp感染与消化性溃疡
从现有资料要确立Hp与消化性溃疡(peptic ulcer diseases,PUD),包括胃溃疡、十二指肠溃疡之间的病因关系尚有困难。部分原因是缺乏合适的动物模型,而且在Hp感染者中仅有少数人发生溃疡。然而几乎所有PUD患者都有Hp感染性胃炎。因此,在没有其他促发因素,如服用甾体类抗炎药(NSAID)或有Zollinger-Ellison综合征存在时,Hp感染与胃溃疡之间的相关性稍弱,不是NSAID诱发的胃溃疡患者中,80%有Hp感染。然而重要的是要注意到Hp感染者中的大多数人并不发生十二指肠或胃溃疡。这些事实意味着宿主的遗传性特征,菌株的变异性或其他因子在其中起着作用。Hp感染根除后,溃疡病复发率显著降低,这是Hp在PUD中起病因作用的最有力的证据。Hp根除后可预防溃疡病复发,这在胃溃疡中证据不如十二指肠溃疡那样充分。在十二指肠溃疡病例中,奇怪的是有些研究中,Hp在胃窦部部位比发生溃疡的十二指肠部位更为常见。胃窦部位的Hp为什么会引起十二指肠病变?已经提出的机理包括:Hp在十二指肠胃化的细胞上定居,胃酸或十二指肠碳酸氢盐的继发性改变,或感染菌的产物和/或宿主炎症应答反应引起的病变。
1.2.3 Hp感染与非溃疡性消化不良
Hp感染与非溃疡性消化不良(NonulcerDyspepsia,NUD)的相关性尚缺乏令人信服的证据。因此尚存在着争论。NUD患者中Hp的感染率并不高于一般人群。
1.2.4 Hp感染与胃部恶性肿瘤
Hp感染与胃腺癌:胃腺癌是世界上最常见的恶性肿瘤之一。在我国发病率仅次于肺癌。有证据显示Hp感染与胃体和胃窦腺癌相关联。然而,胃癌亦发生于一些无Hp感染证据的人。Hp感染的预防和治疗对胃癌发生的危险性的影响,证据尚不够充分。流行病学资料表明:胃癌发生率在一些Hp感染率高的人群中较高。Hp感染率和胃癌发生率都与社会经济状态呈负相关,都随着年龄增加而升高。在发达国家的连续出生列队研究中已见下降。在美国,非洲裔美国人和拉丁美洲人的发生率高于白种人。已发现,Hp感染率与胃癌死亡率之间存在着地区分布性的相关。然而,这两种病在流行病学上存在一些明显不一致的情况。例如,胃癌男人比女人高发,而Hp感染率两性间并无差别。据报道,某些人群Hp感染率高,但胃癌发生率低。这些不一致表明:除Hp感染率外其他因素在胃癌危险因素中亦很重要。在有一些但不是全部回顾性血清学研究中表明,胃癌患者的Hp感染率比对照组高。
感染与胃癌有关的最有力的证据来自三个前瞻性列队血清学研究。这些研究表明Hp感染者的胃癌发生率显著增加。在这些研究中,均未见Hp感染与贲门癌和胃食管联结处癌有关联。
Hp感染与淋巴瘤:胃的非何杰金淋巴瘤是一种罕见的病,占胃恶性肿瘤的3%。粘膜相关淋巴组织性淋巴瘤(MALT)低度克隆的新生物,被认为起源于固有膜中的淋巴样聚合(Lymphoid aggregate)。初步的流行病学资料显示,Hp感染与胃非何杰金淋巴瘤和MALT淋巴瘤都相关。
如果Hp感染与胃癌之间有任何病因关系,那么显然别的因素在胃癌的发生率中亦有重要作用。因此,目前尚不提倡出于预防胃癌的目的而作根除Hp的治疗。
2 Hp的生物学性状
2.1 Hp的形态学特征
Hp的形态学已有了充分的描述。总之,它是一种单极、多鞭毛、末端钝圆、螺旋形弯曲的细菌。(图1)长2.5~4.0μm,宽0.5~1.0μm。革兰染色阴性。有动力。在胃粘膜上皮细胞表面常呈典型的螺旋状或弧形。(图2)在固体培养基上生长时,除典型的形态外,有时可出现杆状或圆球状。
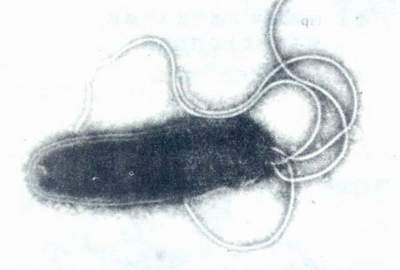
图1 Hp的悬滴负染标本透射电镜(×48000)
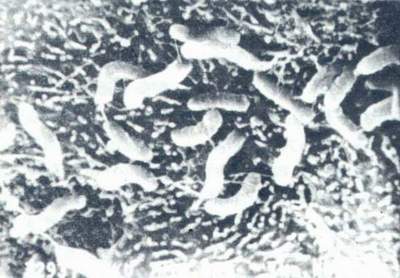
图2 Hp藉鞭毛与部分菌体外膜粘附于胃粘膜上皮细胞表面扫描电镜(×16000)
电镜下,菌体的一端可伸出2~6条带鞘的鞭毛。在分裂时,两端均可见鞭毛。鞭毛长约为菌体1~1.5倍。粗约为30nm。鞭毛的顶端有时可见一球状物,实为鞘的延伸物。每一鞭毛根部均可见一个圆球状根基伸入菌体顶端细胞壁内侧。在其内侧尚有一电子密度降低区域。(图3)。鞭毛在运动中起推进器作用,在定居过程中起抛锚作用(图3)。
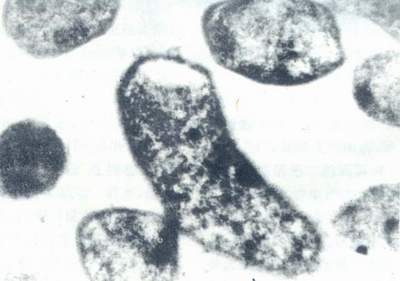
图3 Hp菌体末端及鞭毛根部的超微结构超薄切片透射电镜(×5000)
Hp经鞣酸处理,发现其外表面被厚达40mm的糖萼(glycoculyx)所包裹。电镜下,由于其呈细丝网状,与胃上皮细胞表面连接,因此亦有人称之为纤毛或拟菌毛。它成为Hp粘附于胃上皮细胞表面的主要物质基础(图4)。Hp定居于胃上皮细胞表面,除了鞭毛和糖萼的作用以外,在电镜下,尚可见到菌体细胞壁与胃上皮细胞细胞膜表面直接相贴的现象(图5)。这种现象的机理可能与藉糖萼粘连的机理不一样。其本质及其有何特殊意义有待研究。
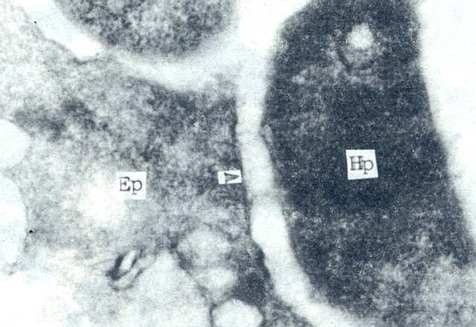
图4 Hp藉拟菌毛粘附定居于胃上皮细胞表面超薄切片透射电镜(×70000)
Hp:幽门螺杆菌
Ep:胃上皮细胞
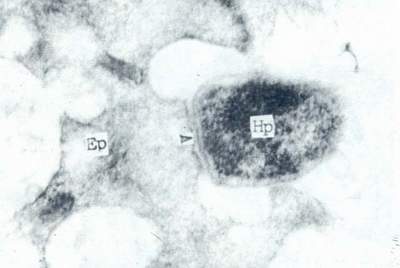
图5 Hp部分细胞壁与胃上皮细胞部分细胞膜直接相贴超薄切片透射电镜(×60000)
Hp:幽门螺杆菌
Ep:胃上皮细胞
培养物中的Hp,用悬滴法作电镜观察时,常见菌体外围有“炸面圈样”或“指突状”结构包裹。这些结构有何含义亦不清楚。
延长培养时间,典型的细菌会发生圆球样形态的变化。这可能就属于Hp的L型。它亦可在暴露于大气(含O2量较高)时发生。这种圆球体(图6)有两种类型。一种较大,电镜下可见稀疏的细胞质,细胞体积膨大。这种类型可能是一种退化型,在传代中不能再生长。另一种小圆球体,电镜下可见电子密度较高的细胞质,且有完整的细胞膜。这种类型可能比前者有较高含量的KDO和蛋白质。它已经被证明至少在30d内对物理和化学因素有一定的耐受性。且在4周~6周内在合适的培养条件下能重新生长成繁殖体。可是迄今尚未能证实这种圆球体在自然环境中的持续存在。或许这是弄清Hp感染的流行病学和某些治疗中失败的原因的关键因素。
2.2 Hp的生理学特征
2.2.1 Hp的生长和生存条件
Hp是一种专性微需O2菌。它的稳定生长需要依靠在生长的微环境中含2%~8%O2。因此,它在大气中和绝对厌O2条件下均不能生长。从临床标本中分离野生株都必需补充适量的CO2。早期认为Hp并不在代谢中利用碳水化合物获得能量,而是利用有机酸和氨基酸。但是不久前Reymolds等研制了一种合成培养基证明精氨酸、组氨酸、异亮氨酸、亮氨酸、甲硫氨酸、苯丙氨酸、缬氨酸是Hp的必需氨本酸。有一些株尚需要丙氨酸或丝氨酸。但没有葡萄糖时,Hp仍不能生长。有适量葡萄糖和丙氨酸时能大大促进其生长。这说明葡萄糖可能仍然是Hp能量和碳源的重要来源之一。Hp只含有MK-6呼吸醌,不含有所有弯曲菌属中普遍存在的甲基化MK-6呼吸醌。
许多固体培养基能用作分离培养Hp的基础培养基。例如:心脑浸液琼脂、哥伦比亚琼脂、布氏琼脂和M-H琼脂等。但必需加入适量的全血(马、羊或人)或胎牛血清作为补充物。加入适量的活性炭(0.2%)或淀粉(1%)亦有利于吸收培养基中衍化产生的毒性氧离子。Hp生长缓慢,通常需要3d~5d,甚至更长时间,才能形成针尖状小菌落,为了避免快速生长的兼性厌O2菌和霉菌等和过度生长,常需加入由万古霉素、TMP、两性霉素、多粘菌素等组合的抑菌剂。Hp在液体培养基中生长更加困难。这可能与在液体里面更难保证菌体周围稳定的微含O2量环境有关。
有人报告Hp能在33°C~40.5°C和pH6.6~8.4条件下生长。但实际应用中仍以37°C和pH7.0~7.2为最适条件。Hp对低ph 比一般细菌有较强耐受力。电镜下,甚至偶然可见Hp钻入壁细胞的分泌小管中(图7)。Clyne等认为酸对pH既有杀伤作用的一面,亦有保护作用的一面。他们把尿素酶阳性的野生株N6和尿素酶阴性的突变株分别接种在含有或没有10mmol尿素的两个pH范围从2.2到7.2的37°CPBS中,30min,然后测量CFU/ml、上清液的pH和溶液中的氨量。结果N6株在没有尿素的pH4.5~7.0的PBS中生存良好,甚至在有尿素的超始pH为3.5的溶液中亦能生存。然而野生株和突变株均不能在碱性环境中生长繁殖。存在野生株和尿素的酸性溶液的pH迅速从3.5升至8.45。尿素酶突变株在pH4.5~7.2的溶液中存活,不对尿素起反应。野生株N6暴露于氨的浓度高达80mmol仍能存活。胃的酸性环境可能对存活于尿素中的Hp有杀伤作用。但是Hp决不能存活于含有尿素的正常环境中,因为随后升高的pH的危害性远超过氨的毒性。
在血琼脂中,Hp不能在含有5%胆汁的情况下生长。但是暴露于含有5%胆汁的液体培养基中30min,只有25%的Hp被杀死。这说明Hp通过十二指肠时尚有生存的可能性。
在含有0.1 %到1.5%甲基纤维素的1%蛋白胨的粘稠深液中,有周鞭毛的大肠杆菌在20CP(厘泊),即被制止运动。而Hp在10CP比1CP游动得更快,甚至在200CP的粘稠培养基中仍能游动。这可能与Hp的螺旋形特征及鞭毛的旋转推动作用有关。由此,使Hp很容易穿透胃粘膜表面的粘液层,达到胃粘膜上皮细胞表面定居。而其他细菌很难达到这一点。
2.2.2 Hp的生化反应和酶
Hp对临床实验室中常用于鉴定细菌的大多数经典试验不起反应。但是有几种酶的反应是有特点的,特别是尿素酶,常用作Hp的生化鉴定。
尿素酶 尿素酶是Hp的一个非常重要的酶。它能使尿素分解成二氧化碳与氨。用柱层析方法测得其分子量约为625±kD,与底物有高度亲和性。它与变形杆菌属、摩根菌属、普罗威登斯菌属等许多微生物产生的尿素酶相比,在分子量大小、电泳活性、等电点和对尿素酶抑制剂的敏感性等方面都有它的独特性。Hp在固体培养基上和胃粘膜上能产生大量尿素酶,而且其酶的活性比其他微生物产生的尿素酶活性高几倍到几十倍,甚至更大。尿素酶不仅是Hp得以在胃粘膜表面定居的必要条件,可能也是Hp在胃部致病的重要因素之一。传统的测定尿素酶的方法,如Christensen尿素培养基等,均可用于测定Hp的尿素酶。由于Hp的发现和临床上对快速诊断Hp感染的需要,已由此发展出了许多种更加简便有效的或者能作非侵袭性体内诊断用的试验。
其他可用于Hp常规鉴定用的酶试验Hp的分离培养物的生化特性往往非常稳定。它们都能产生氧化酶、触酶、尿素酶、碱性磷酸酶、α-谷氨酰胺肽酶、亮氨酸胺肽酶和DNA酶。不产生其他44种酶(包括许多种氨肽酶、酯酶、糖甙酶等)。常规鉴定可参见表1。
表1Hp与相关细菌的生化特征比较
| Hp | Hf | Hm | Ws | Cja | |
| 尿素酶(快速) | + | + | + | - | - |
| 氧化酶 | + | + | + | - | + |
| 触酶 | + | + | + | + | + |
| H2S产生 | - | - | - | - | - |
| G+C mol% | 37 | 42.5 | 36 | 47 | 30-38 |
| 形态 | 弧形或螺形 | 紧密螺旋 | 直到弧形 | 短弧 | 短弧 |
| 硝酸盐还原 | - | + | + | + | + |
| 马尿酸水解 | - | - | - | - | + |
| 碱性磷酸酶 | + | + | + | - | - |
| 精氨酸胺肽酶 | + | + | + | + | + |
| 组氨酸胺肽酶 | + | + | + | - | - |
| 亮氨酸胺肽酶 | + | + | + | + | + |
| γ-谷氨酰转肽酶 | + | + | + | - | + |
| 萘啶酸耐药(30μg/ml) | + | + | - | + | - |
| Cephalothin(30μg/ml) | - | - | - | + | - |
| 含1%甘油生长 | - | - | - | - | + |
| 含1.5NaCl生长 | - | - | - | -NDb | + |
| 42°C生长 | - | + | + | +/- | + |
| 37°C生长 | + | + | + | + | + |
| 25°C生长 | - | - | - | - | - |
aHp=Helicobacter pylori Ws:Wollinellasuccinegenes Hf=Helicobacter Cj:Campylobacter jejuni Hm=HelicobactermustelaeNDb:未确定
2.2.3 对抗菌药物的敏感性
在体外,Hp对大多数抗菌制剂敏感。但对万古霉素和TMP高度耐药。磺吡苄头孢霉素(Cefsulodin),多粘菌素B及萘啶酸对少数菌株以外也都不起作用。这些抗菌药物可用于制备Hp的选择性培养基。许多抗菌药物在体外对Hp的高敏感性和在体内的低效或无效之间的矛盾,可能涉及许多因素。除了Hp自身发生的耐药性突变外,恐怕还受到药物在胃腔内对Hp作用的条件(胃酸作用的强度、不溶性粘液层的阻隔、不断的排空运动等)影响。Hp对常用抗菌药物的敏感性见表2。近年来由于临床上广泛应用灭滴灵、克拉霉素(Clarithromycin)等抗菌药物治疗慢性胃炎及消化性溃疡。因此,临床上对这些药物的耐药菌株已越来越多见。
表2 Hp对常用27种抗菌制剂的敏感性
| MIC(μg/ml)a | |||
| 范围 | 50% | 90% | |
| 青霉素G | 0.015~0.12 | 0.06 | 0.12 |
| 氨苄青霉素 | <0.003~0.03 | 0.015 | 0.03 |
| 克拉维酸 | <0.01~0.64 | 0.16 | 0.64 |
| 羟氨苄青霉素+克拉维酸 | <0.01~0.02 | <0.01 | 0.01 |
| 先锋霉素Ⅰ | 0.025~0.4 | 0.2 | 0.2 |
| 氨噻肟头孢霉素 | 0.01~0.16 | 0.04 | 0.08 |
| 磺毗苄头孢霉素 | 5.12~41.0 | 20.5 | 41.0 |
| 先锋霉素V | 0.2 | 25.0 | |
| 链霉素 | 0.04~1.28 | 0.32 | 0.64 |
| 卡那霉素 | 0.04~0.64 | 0.16 | 0.32 |
| 妥布霉素 | 0.04~0.64 | 0.08 | 0.16 |
| 庆大霉素 | 0.04~0.32 | 0.08 | 0.16 |
| 红霉素 | 0.1~0.8 | 0.2 | 0.4 |
| 交沙霉素 | 0.4~1.6 | 0.8 | 0.8 |
| 洁霉素 | 3.2~12.8 | 6.4 | 12.8 |
| 氯霉素 | 2.0~8.0 | 2.0 | 4.0 |
| 四环素 | 0.01~0.16 | 0.08 | 0.16 |
| 利福平 | 0.5~2.0 | 1.0 | 1.0 |
| 哌氟喹酸 | 1.0~8.0 | 4.0 | 8.0 |
| 多粘菌素B | 6.25 | 50.0 | |
| 多粘菌素E | 2.0~64.0 | 8.0 | 32.0 |
| 万古霉素 | 50.0~>100 | >100 | >100 |
| TMP | >100 | >100 | |
| 次硝酸铋 | 3.12 | 25.0 | |
| 灭滴灵 | 1.56 | >100 | |
| 痢特灵 | <0.05 | 0.2 | |
| 克拉霉素 | 0.05 | 0.65 | |
aMIC:Minimal inhibitory concentration
2.2.4 粘附性
自然条件下,大量Hp仅出现在胃上皮细胞表面,细胞间隙及胃小凹中,而不出现在其他组织细胞表面(图8),说明Hp对胃上皮细胞有特殊的亲和性。后来人们也发现Hp对某些动物和人的RBC会发生聚集反应。作者还在Hp与“O”型血的血凝块的超薄切片中发现,Hp与“O”型RBC(后来证实按Lewis分型属Leb型”,及Hp与胃粘膜上皮细胞之间的关系,在高倍电镜下所见非常相似(图9)。这些特异的亲和现象很可能暗示着类似的粘附机制。1993年有人发现Hp特异地粘附于人胃粘膜上皮细胞受到人类初乳中SCIgA(一种携带高度多变的N-和O-连接的单糖链的糖蛋白)的抑制。当这种SC-IgA被墨角藻糖酶(α-L-fuco-sidase)降解后,抑制作用即降低,若用唾液酸(sialic acid)处理(曾有人报道Hp表达唾液酸特异的凝集素),并不降低其结合力。这一结果证明Hp在胃粘膜上皮细胞上的受体含有墨角藻糖(fucose)。而墨角藻糖结合的血型抗原,典型地在RBC上发现,也在人上皮细胞表面表达。Boren等为了弄清墨角藻糖酶敏感的Hp受体结构,他们比较了sc-IgA与S-IgA分子含有的不同墨角藻糖碳水化合物链的棋盘分析(Panel analysis)。最后他们证实了在人胃粘膜表面的Hp受体含有RBc Lewis分型的Leb抗原。他们还证实了Leb抗原是分泌型体质个体胃上皮细胞表面表达的最显著的血型相关抗原,而Lea 抗原是非分泌型体质个体中占相当优势的血型抗原。因此,阳性分泌型体质个体可能对Hp比较易感。胃溃疡个体的流行病学调查中也证明了Hp感染在Leb阳性的个体中较高发。
2.2.5 菌种保存
传统的冻干方法并不适宜于Hp的保存。有人曾用棉签沾取Hp的液态培养物冻干取得成功。但是目前置-70°C或置液氮中冷冻是常用保存方法。
2.3 Hp的分子生物学特征及菌株标识
2.3.1 分子生物特征
脂肪酸组成,一种细菌独特的脂肪酸组成常常与该菌的分类命名、细胞膜的生理化学特性和该菌的生物学性状有关。在早先许多学者都对Hp的脂肪酸组成作了测定。尽管测得的数值略有出入,但多数学者认为Hp经甲基化处理由气相色谱仪测得的数值表明:Hp的主要脂肪酸有十四烷酸(C14:0)、十六烷酸(C16:0)、十六碳稀酸(C16:1)、十八烷酸(C18:1)、顺式9、10亚甲基十八烷酸(C19:0cyc),但是缺少了一羟基十四烷酸(3-OH-C14:0)。
Inamoto等深入地研究了Hp的脂质和脂肪酸。他们发现Hp的脂肪主要由胆固醇酯、三油酸甘油酯、游离脂肪酸、胆固酸、二酰化甘油和一酰化甘油等简单脂质组成。且在这些脂质中均含有11-甲氧基十七烷酸(11-OMeC17:0)和11-甲氧基十九烷酸(11-OMe C19:0)两种独特脂肪酸。与BCG的耐酸性相比,两者均有耐酸性,虽程度有所不同。他们认为Hp脂质与脂肪酸的特征可能是构成Hp亦具有耐酸性和与BCG耐酸强度不同的基础。
蛋白质组成Hp的菌体蛋白质含量通常用SDS-PAGE方法测定。用这一技术测得的结果,不同分子量的蛋白质条带很多,不同菌株看上去似乎很一致。但是用激光光密度计扫描,并用电脑分析比较,显示它们之间是存在差别的。所有的株的蛋白质条带80%是相似的,若以91%相似为阈值,可把Hp分成不同的电泳型。Hp的SDS-PAGE电泳谱中部分条带含有Hp的特异性抗原,也有一些条带含有与弯曲菌属细菌的共同抗原。由于技术条件未统一,各家报道的结果均略有上下。1990年Cover等从消化性溃疡患者活检标本中分离到的Hp,在肉汤培养物中用SDS-PAGE测到一种82kD大分子量的蛋白质。它只能在使细胞产生空斑作用的上清液中出现。另外用免疫印迹法以人血清能在产生空斑作用的上清液中识别出128kD的蛋白质条带。两者在产生空斑作用的上清液中出现的频率之间没有显著性差异。但是由于82kD蛋白质不易被人的血清识别。而对128kD蛋白质起血清学反应的抗体在消化性溃疡病人中比无消化性溃疡病的Hp感染者中更为常见。这意味着这两种大分子量蛋白质可能与Hp的致病性有关。后来人们把前者82kD蛋白质称作空泡毒素CacA,把后者128kD蛋白质称作细胞毒素相关蛋白CagA。而且经其他学者的工作发现,CagA蛋白质的分子量飘移在128-140kD之间。项兆英等人对43株Hp CagA和VacA毒力因子表达的分析揭示,能把临床分离株分成两种主要类型。Ⅰ型细菌含有CagA和VacA基因,表达CagA和VacA蛋白;Ⅱ型细菌不含CagA基因;不表达CagA和VacA蛋白。Ⅰ型和Ⅱ型细菌分别占56%和16%,而其余的为中间表型,即仅表达其中一种毒力因子。这一发现证明尽管许多细胞毒性Hp株含有CagA,但是VacA的表达可以不需要CagA的存在。
Hg的鞭毛可将其液体培养物藉着震荡脱落下来,然后再经过梯度离心等方法使其纯化。现知鞭毛蛋白中含有57kD与56kD两种鞭毛素亚单位,它们与弯曲菌属的细菌的鞭毛素具有共同的抗原决定簇。但是56kD的鞭毛素尚具有对Hp特异的氨基酸序列结构。
核酸 早期人们企图判断Hp是否与弯曲菌属同属,测试了它的DNAG+Cmol%,结果发现其数值与弯曲菌属的数值范围(30mol~38mol%)完全重叠。因此这一方法在鉴别Hp与弯曲菌上毫无意义。后来人们根据Hp的16SrRNA才发现与弯曲菌属细菌不同。随着分子生物学的发展,人们迅速地投入了对Hp各种特殊的基因的研究。最早克隆成功的是尿素酶基因。到1995年7月EHpSG(EuropeanHelicobacter pylori Study Group)在爱尔兰召开的第八届胃十二指肠病理学与幽门螺杆菌国际会议上已介绍了不下十余种Hp不同的基因组或基因的研究工作。基中包括:趋化因子cheA和cheY、鞭毛素基因flaA和flaB、鞭毛生物合成调节基因flbA、鞭毛外鞘蛋白基因、转运系统基因nixA、热休克蛋白质基因hspA、尿素酶基因ureA、ureB、ureC、修复基因racA、空泡毒素基因VacA、细胞毒素相关基因CagA、CagC,细胞毒素第二相关基因CagII碱性磷酸酶基因组等。由此可见,对一种病原性细菌来讲,Hp基因研究所涉及领域之宽,进展速度之快是前所未有的。为了避免在这一研究领域发生混乱,大会上有一些学者呼吁为Hp的基因和基因组的统一命名制订规则。他们建议:①Hp基因同源于其他细菌中已经发现的基因,给于同样名称。例如ureA、ureB、flaA等。②完全新发现的基因,根据它们编码的蛋白质的功能或作用命名。例如:VacA。③当同源性不是太明显,新基因的功能还未完全确立,可以给予一个独特的临时性的名称。例如:CagA、CagII等。下面就研究得较多和较重要的两类基因作一介绍。
①尿素酶基因:1988年Labigne用穿梭载体(shurttle vecter)Pill500把Hp对应于尿素酶基因的DNA片段克隆出来,在大肠杆菌和空肠弯曲菌之间复制和转移获得了成功。但是它并不能在大肠杆菌中表达尿素酶活性,只有在接合于空肠弯曲菌时才能暂时地生物合成尿素酶。重组的粘尾质粒(recombinant cosmid)piLL585,具有33.2kb。它经过再克隆成8.1kb片段piLL590后才能把尿素酶的表型特征转输给空肠弯曲菌的受体株。此后,经过反复不断再克隆删除了不必要的部分后,尿素酶的基因最后定位在DNA的4.2kb区段内。用双脱氧法(dideoxy)测序,发现有四个开放读框(open reading frame),分别编码四个预知分子量的多肽。它们分别是26000(ureA)、61600(ureB)、49200(ureC)和15000(ureD)。ureA和ureB编码的多肽与尿素酶结构的两个亚单位是相当的。它们与奇异变形杆菌尿素酶二个亚单位和刀豆尿素酶独一无二的亚单位高度同源。符合率分别是56%和55.5%。虽然ureD编码的多肽,与膜穿透蛋白质功能有关。但是对这一多肽与ureC编码的多肽均未明确其作用。从DNA序列图谱上看,表明这些多肽对把尿素酶活性转移给空肠弯曲菌受体是必需的。
②细胞毒素相关基因CagA和空泡毒素VacA:自从发现Hp高分子量抗原与消化性溃疡病有关后,引起了人们对它的基因的兴趣。Tumuru等用λZapII构建了Hp84-183的随机染色体片段文库。在大肠杆菌YLi-Blue的多个文库中用Hp感染者经过吸收的血清筛选。结果分离纯化出一个3.5kb插入片段的克隆。这一插入片段的再克隆能表达出一种Hp的重组蛋白质。它的分子量接近96kD,且能被人的血清所识别。得自Hp感染者和能识别天然的120~128kdHp抗原的血清,它们识别重组的96kD PMC3蛋白质的能力明显地大于不能识别天然的Hp抗原的血清。所有19株Hp产生的120kD~128kD抗原均能与PMC3产生斑点杂交。没有一个不产生同一抗原的得到同一结果。(P<0.001)。因为15个产生空泡毒素的Hp临床分离株均与PMC3产生杂交。Tumuru等称这一基因为细胞毒素相关基因(cytotoxin-associated gene,CagA)。PMC3的序列分析证实有一个含859个氨基酸的开放读框,没有终端子。另外用人血清筛选λgtII文库还显示出一个能产生噬菌斑的0.6kb插入片段,其序列与PMC3下游序列吻合。为了克隆出全长度的基因,他们用0.6kb片段为探针从λZapII基因库中分离出一个含2.7kb插入片段的克隆。这一插入片段pYB2的核苷酸序列显示为785bp,与pMC3下游区域的序列恰好重叠。CagA全核苷酸序列的翻译揭示有一个产生分子量达131.517kD的1181个氨基酸组成的蛋白质的开放读框。这些结果与先前报道过的蛋白质序列没有明显的同源性。这些发现表明Hp高分子量抗原的克隆和特征可能与Hp的毒力和空泡毒素的产生有关。
至于Hp的空泡毒素,Telford根据已知Hp空泡毒素23个氨基酸组成,取其分别对应于最初和末了6个氨基酸组成的单核苷酸序列作PCR反应。并以PCR产物作探针探查Hp全菌DNA用HindIII酶切的文库,并克隆入其本质粒载体(bluescript plasmidvector),分离到含有~3kb插入片段的克隆。随后从用EcoRI酶切的文库分离出的7kb插入片段,也覆盖HindIII克隆的120bp区段。这两个片段均含有完整的基因。此基因含有唯一的较长的开放读框,能编码1296个氨基酸,分子量为139.7kD的蛋白质。由起动子,核糖体结合部位,终端子组成。最前面的33个氨基酸似乎是细菌信号肽,提示蛋白质是藉分泌决定机制(Sec-dependent mechanism)输出的。从34~56序位是与纯化的NH2-终端编序决定的23个氨基酸是一致的。然后,他们又用基因的不同区段在大肠杆菌中表达,产生了五种融合蛋白。并用免疫印迹法最后证实94kD的融合蛋白与前述的空泡推测基因相对应。
2.3.2 菌株标识
有人曾经企图对Hp进行生物学分型,但是没有获得明显的成功。尽管对蛋白质的SDS-PAGE及对全菌DNA的指纹分析会显示菌株之间的微细差异,但似乎并不实用。因为菌株与菌株之间的差异太微细、太多,难于识别。近年来在PCR反应发展的基础上出现了RAPD(Randomamplified polymorphic DNA)和RFLP(Restriction fragment length polymorphism)两种识别Hp不同种株的技术,这将为Hp的流行病学调查提供一些有效的方法。
3 Hp的致病与免疫性
Hp感染几乎总是与炎症反应同时并存。但是消化性溃疡与胃癌只在一小部分Hp感染者中发生。尽管国内外对Hp感染的致病机制已从临床资料及体外试验中作了大量资料积累。对它的致病机制也有了一些初步解释。但是仍然很少经过合适的动物模型的验证。因此它的确切致病机制现在仍然知道得不是很多。至于它的免疫性更是如此。
3.1 Hp的致病性
下面就胃十二指肠几个与Hp感染有关的主要疾病的发生学说作一些介绍。
3.1.1 胃炎的发生学说
正常情况下,胃壁有一系列完善的自我保护机制(胃酸、蛋白酶的分泌功能,不溶性与可溶性粘液层的保护作用,有规律的运动……等),能抵御经口而入的千百种微生物的侵袭。自从在胃粘膜上皮细胞表面发现了Hp以后,才认识到Hp几乎是能够突破这一天然屏障的唯一元凶。尽管也有报道另一种人胃螺菌(Gastrospirilumhominis,Gh,现又有人称之为Helicobacter heilmanii)亦能在人胃壁上定居,但其阳性检出率所占比例平均一般仅及Hp的1%以下。Goodwin 把Hp对胃粘膜屏障在破坏作用比喻作对“屋顶”的破坏给屋内造成灾难那样的后果,故称为“屋漏”学说。目前对Hp感染的研究能归入这一学说的资料最多(见表3)。主要包括①使Hp穿透粘液层在胃上皮细胞表面定居的因素;②对胃上皮细胞等起破坏作用的毒素因子;③各种炎症细胞及炎症介质;④免疫反应物质等。这些因素构成Hp感染的基本病理变化,即各种类型的急、慢性胃炎。其中近年来得到最重要关注的是空泡毒素VacA、细胞毒素相关蛋白质CagA,和尿素酶等的作用及其分子生物学研究(见前述)。
表3 Hp可能的致病因子及炎症反应
| 致病因子 | 可能的作用 | |
| 定居及毒性因子 | 螺旋形菌体结构 | 有利通过胃粘液层 |
| 鞭毛 | 加速穿透粘液层 | |
| 粘液素酶 | 降解糖蛋白、有利Hp穿透粘液层 | |
| 尿素酶 | 降糖尿素,既有利Hp生存,又对上皮细胞有毒 | |
| 粘附素 | 使Hp定居胃上皮细胞表面 | |
| 蛋白酶 | 降解粘液层,破坏上皮细胞膜 | |
| 脂酶和磷脂酶A | 消化粘液层,破坏上皮细胞膜 | |
| 过氧化氢酶 | 在胃粘膜上起抗氧化作用,保护Hp自身 | |
| 空泡毒素VacA | 破坏上皮细胞,使胞内物质溢出 | |
| CagA蛋白质 | 作用未明 | |
| 胃酸分泌物抑制因子 | 有利急性炎症发生 | |
| 脂多糖 | 能抑制上皮细胞膜基质的合成 | |
| 炎症反应 | 粘液层完整性破坏 | |
| 中性粒细胞的激活 | ||
| 单核细胞与巨噬细胞的激活 | ||
| 白三烯B4(ITB4)的继发性增加,增强炎症 | ||
| 白细胞移动抑制因子的作用 | ||
| 磷脂酶的降解物的趋化和改变细胞膜通透性的作用 | ||
| 血小板激活因子促进局部粘膜的损坏 | ||
| 65kD热休克蛋白与胃窦上皮有交叉抗原作用 | ||
| 浆细胞分泌的IgA激活酸性粒细胞降解破坏粘膜固有层 | ||
| 碱性粒细胞释放组织胺,与IgE结合释放血管活性因子 |
3.1.2 消化性溃疡的发生学说
Hp感染明显地增加了发生十二指肠和胃溃疡的危险性。大约1/6Hp感染者可能发生消化性溃疡病。治疗Hp感染可加速溃疡的愈合和大大降低溃疡的复发率。不用抑酸剂,单用抗Hp药物治疗,表明也能有效地治愈胃和十二指肠溃疡。Hp感染已经与一些引起溃疡病的原因找到了联系。例如:胃酸增加、十二指肠胃化生、粘膜屏障性质的改变、胃窦粘膜产生炎症代谢产物等。这些患者中的发现已在动物实验中得到初步证明。实际上消化性溃疡涉及几个复杂的相互作用的机制。如细菌的毒力因素(VacA和CagA等),宿主的反应性(例:如易感性的遗传、十二指肠上皮的胃化生、粘膜屏障和炎症的相互作用、泌酸反应、神经调节作用)和环境因素(例如饮食、获得感染的年龄)的综合作用导致溃疡的最后结果。过去临床上对溃疡的发生有一句谚语,叫“no acid,no ulcer”。现在,从现代理论来看,“no Hp,noulcer”应得到更多地强调。
3.1.3 胃癌的发生学说
胃癌是世界上肿瘤中仅次于肺癌的死亡原因。美国胃癌于1930年占肿瘤死亡原因首位。到现在已下降到较低水平(1985年为第9-10位)这一发生率的显著下降有利于研究这一疾病的致病机制.事实上,作为结果,胃癌可被认为在人类恶性病变中了解最清楚的。自从发现了Hp以后,对胃癌的发生提出了一些新的看法。
从早期描述的流行病学和临床资料研究的结合,肠型胃癌的学说早已形成。当时认为环境因素导致胃刺激,引起浅表性和萎缩性胃炎。低胃酸症,接着发生细菌过量生长,使亚硝酸盐转化成N-硝基胺(N-nitrosamines),引起组织异化,最后发生肿瘤。维生素C和β-胡萝卜素藉着它的抗硝基胺作用和抗氧化物作用会防止疾病的后期发展。这一得到广泛支持的学说是很有力的,但不能解释所有问题。自从清楚Hp感染是慢性胃炎主要原因,且在未治疗条件下几乎可以持续终生,引起了人们对胃癌形成学说的修正。从近年来对Hp感染的大量研究中提出了许多Hp致胃癌的可能机制:①细菌的代谢产物直接转化粘膜;②类同于病毒的致病机制,Hp DNA的某些片段转移入宿主细胞,引起转化;③Hp引起炎症反应,其本身具有基因毒性作用。在这些机制中,后者似乎与最广泛的资料是一致的。
体外和体内实验中,均见Hp引起细胞增殖。有人提出这是淋巴因子中的上皮细胞生长因子引起的。另一些人把它与尿素酶产生的氨联系起来。炎症细胞的增加对上皮细胞的增殖会增进危险性。中性粒细胞产生O2、H2O2、HOCL和氯胺等都能引起DNA键断裂,损害碱基对,和姊妹染色单体交换(sister chromatidexchanges)。突变原性的反应性O2产物的过量产生也已被Hp感染人类粘膜组织的化学发光(chemiluminesence)所证实,由此,Hp感染不仅引起有丝分裂,还提供了内源性突变原的来源。
Hp感染引起的细胞增殖,藉着①复制错误,②内源性炎症相关突变原,③饮食中外源性突变原三种机制增加对DNA损坏的危险性。大多数损坏的DNA能被机体正常的保护机制所修复。但是检查和修复的能力并不是总是那样的完美无缺的。上皮细胞中引起的有些DAN损坏可保留很长时间。感染期越长,不适当的修复可能性越大。最后转化成恶性。人们感染Hp在年青时期,他们产生明显的炎症反应,会增加肿瘤的危险性。假若这些人包含中会有较高的外源性致癌原和较低的抗氧化剂,它们的危险性会互加起来。虽然有些个体中感染或饮食已能单独解释肿瘤的成因,但两者的互加更能显示它们之间的协同作用。
3.2 Hp的免疫性
3.2.1 对Hp的体液免疫反应
通常对大多数Hp感染体液免疫反应的研究只是用在诊断上。Doig等用单克隆抗体的方法描述了Hp表面暴露抗原蛋白质的性质。他们鉴定的暴露表面的抗原包括有分子量分别为80、60、51、50、48、和31kD的各种外膜蛋白质。LPS的核心区域和鞭毛外鞘也被识别为Hp抗原。50、48和31kD的是不同株所共有的,而其余的为株特异性抗原。这些抗原成分和在感染病人血清中相应的抗体可能是有用的致病性和诊断的标记。用免疫印迹法,不同感染病人血清中并不能测得同样的抗原组成。这提示病人对这些抗原的抗体反应的个体差异。而且这提出了一个问题:这种抗体反应异质性的意义是什么?
Krenning等研究了IgG、IgA和Hp定居密度及胃炎严重程度的关系,他们认为IgG反应的强度与Hp定居的密度及胃炎的严重程度相关。但是IgA反应与胃窦炎强度有关,而不与胃窦部Hp定居密度和活性程度有关。IgA水平也与胃体部Hp定居密度和胃炎强度有关。
3.2.2 对Hp感染的细胞和炎症局部反应
在体外受到Hp抗原刺激后,来自Hp阳性病人外周血或胃粘膜固有层的T淋巴细胞的增殖和α-干扰素的产生比Hp阴性病人均有所下降。在Hp阳性病人胃粘膜固有层中某些淋巴细胞亚群(CD8和CD22)比Hp阴性病人中有所增加。这表明活跃的T淋巴细胞受到Hp抗原抑制,且可以部分解释免疫反应为什么对根除Hp无能。
Takemura等为了确定Hp水浸出液激活的中性粒细胞是否会破坏内皮细胞单层,并涉及这一过程的机制。他们把人类中性粒细胞与加入或没有加入Hp浸出液的人类脐带静脉内皮细胞(HUVECs)单层一起培养。结果Hp浸出液激活的人类中性粒细胞使内皮细胞从HUVECs单层上脱落下来。其严重程度取决于暴露的时间,但是内皮细胞的脱落可以藉直接抗中性粒细胞上的CD11/CD18单克隆抗体或抗内皮细胞上的细胞粘附因子1单克隆抗体防止。HUVEC单层的互解也可以藉超氧化物歧化酶、触酶和抗弹性硬蛋白酶(elastase)的单克隆抗体防止。进一步的研究表明Hp的浸出液是能抑制人的中性粒细胞弹性蛋白酶的。抗弹性使蛋白酶的作用不因氧化剂减弱。这些研究表明Hp浸出液激活的人类中性粒细胞能破坏HUVEC单层只有在人中性粒细胞能破坏HUVECs时,由此,对体内观察到的由Hp浸出液引起的中性粒细胞依赖的血管蛋白质泄漏提供了一个解释。Hp释放的多种抗蛋白质酶的可能性,可以部分地解释为什么这一细菌的毒力是如此的强。
4 Hp感染的诊断、治疗和流行病学
4.1 Hp感染的诊断
自1983年通过胃镜取活检标本分离培养成功以来,对Hp感染的诊断已发展出了许多方法,包括有细菌学、病理学、血清学、同位素示踪、分子生物学等许多方法。但总的讲来,从标本采集角度看,可以分为侵袭性和非侵袭性两大类。
4.1.1 Hp感染的诊断
侵袭性方法主要指必需通过胃镜取活检标本检查的方法。它是目前消化病学科的常规方法。它包括细菌的分离培养和直接涂片、快速尿素酶试验,药敏试验。病理学的组织切片检查,电镜检查及分子生物学的PCR,RAPD,RFLP等。其中分离培养+直接涂片镜检或分离培养+病理切征检查被国际上分认为判断其他各种方法准确性可靠性的依据,即所谓“金标准”。可是Hp是一种微需O2菌,分离培养较困难。需要有一定的技术设备条件,因此在许多医院,甚至在较大的医院,亦未能普遍开展此项工作。现在看来,分离培养条件掌握得好,若能在胃壁不同病灶部位(因Hp感染灶在胃壁上分布不匀)取两块活检标本作分离培养,Hp的阳性检出率几乎可以达到90%以上,甚至接近100%。目前临床普遍采用的还是病理组织切片和快速尿素酶两种方法。病理切片HE染色特异性较差,Warthin-Starry镀银染色法较好。快速尿素酶试验由于市场上有药盒供应,有的自己配制,使用很方便。但是由于产品质量标准不统一,结果出入很大,因此在临床应用上造成了一些混乱。
PRC从理论上讲,本来是一种很敏感的、特异性亦较好的方法。但是必须具有合适的引物,良好的设备,严格的实验条件才能成功地进行。因此目前在临床上仅用于研究,尚未普遍开展。随机扩增多形性DNA(Random amplified polymorphic DNA,RAPD)及限制性片段长度多形性(Restriction fragment length polymorphism,RFLP)试验是PCR技术基础上发展起来的两种方法。它们共同特点是能区别Hp株的特异性。因此在追踪Hp的传染源或判断溃疡病的复发是由于Hp未根除还是再感染的有效工具。成熟的PCR技术除了可应用胃及口腔等部位的消化道标本的检测外,还可用于体外标本,为解决Hp的传播途径等问题作出贡献。
这一大类方法最大的缺点,或者说受到的限制是必需通过胃镜取标本。除了消毒不严密,胃镜检查本身就是一种传播Hp感染的媒介以外,胃镜检查还能造成胃病患者额外增加一些损伤与痛苦。一般来讲患者都是在迫不得已的情况下才接受这一检查的。因此更不宜在短期内重复用于抗Hp疗效的考核上。而且做胃镜的费用亦较贵,若不是为了初诊的需要,只是为了考核抗Hp疗效,普遍再去做胃镜检查,显然是不适宜的。
4.1.2非侵袭性诊断方法
国际上是指非通过胃镜取活检标本诊断Hp标本感染的方法.这类方法包括血清学和同位素踪两大类。
血清学方法 是根据Hp感染者血清中出现相应的抗体而设计的。血清中的IgM与IgA亦均能检测到,但其临床意义均不如IgG明显。因此目前最常用的是以ELISA方法测血清中的IgG,或者再加上IgA。这在市场上已有药盒供应。由于各家厂商所用的抗原种类不同。所定的标准又不统一,因此结果参差不齐。若用全菌或粗制抗原,由于与空肠弯曲菌存在共同抗原,敏感性可能较高,但特异性较差。若用第三代精制抗原(测Hp的尿素酶或外膜蛋白等)特异性较高,但敏感性可能会削弱一些。由于患者抗体一旦产生后,会持续相当长时间。因此Hp的血清学反应阳性表明此人必定感染过Hp,或者此人目前还可能在感染中。但是由于胃中Hp经治疗后已被杀灭,血清中IgG不可能一下子降下来,还可持续6~12个月,因此血清抗体测定一般不适宜用作抗Hp疗效的监测。
采取胃液用ELISA法测分泌型IgA,亦是一种可采用的血清学方法。但其实际意义未见有更大优越性,因此未获得广泛采用。最近国外还有人提出用ELISA法测唾液中IgA。若后者方法上趋于成熟,结果可靠,将是一种较有前途的方法。
同位素踪方法 这一类方法是基于Hp尿素酶能把尿素分解成CO2和NH3,根据用不同的同位素标记在尿素的C原子或N原子上,然后让被试者口服一定量的标记尿素,定时收集呼出的气体或排出的尿,检测其中标记CO2和NH3的排出率,即可准确地反映Hp在胃中的存在。由此,产生了三种不同的试验。用13C标记的称作13CO2,呼气试验,用14C标记的14CO2呼气试验。这两种试验方法与作用类同,只是13C只是稳定性同位素,检测仪器需用质谱仪,但目前已可用光谱仪检测。14C是放射性同位素,可用液体闪烁扫描仪检测。14C是一种半衰期长达5000余年的放射性同位素。对被检测者有可能造成长期的内照射损伤,是一种不宜推广应用的方法。更不宜用于孕妇及儿童,在成人中重复应用亦应极其谨慎。1992年我国学者吴继琼等人在国际上首创了用稳定性同位素15N标记的尿素作15NH4+排出试验(后又改称15N排出试验)。这一种试验原理与作用与13CO2呼气试验相同,无放射性损伤。所不同的是搜集的标本不同,前者搜集的是尿样,后者搜集的是呼出气体。目前国际上普遍认为13CO2呼气试验是监测抗Hp疗效的最佳方法。其优点:可无损伤地及时地反映胃内的实际感染情况,敏感性特异性较高,可克服活检标本因Hp感染灶在胃壁分布不匀可能造成的假阴性结果,甚至孕妇儿童都能重复使用。尿15N排出试验与13CO2呼气试验一样,目前由于涉及设备与费用总是,暂时还难于推广。
4.2 Hp感染的治疗
Hp感染现在主要靠抗Hp药物进行治疗。尽管Hp在体外对许多抗菌药物都很敏感,但是在体内用药并不那样如意。这是因为Hp主要寄生在粘液层下面,胃上皮细胞表面。注射途径用药,对它无作用,经口局部又因为胃酸环境.粘液层的屏障及胃的不断排空作用,使药效也大大地受到了限制。再加上有些药长期应用易产生严重的副作用或耐药菌株等问题。因此Hp感染引起的急慢性胃炎.消化性溃疡等疾病,本来看起来很容易治疗的问题,实际上效果并不总是很理想。何况目前缺乏合适的Hp感染的动物模型,可供帮助制订有效的治疗方案。因此目前的治疗方案几乎全凭临床经验制订,有很大的局限性(因地区、人群的差异)。总的讲来,目前不提倡用单一的抗菌药物,因为它的治愈率较低,一般<20%,且易产生耐药性。
治疗方案的选择原则是:①采用联合用药方法;②Hp的根除率>80%,最好在90%以上;③无明显副作用,病人耐受性好;④病人经济上可承受性。判断Hp感染的治疗效果应根据Hp的根除率,而不是清除率。根除是指治疗终止后至少在一个月后,通过细菌学、病理组织学或同位素示踪方法证实无细菌生长。
目前国内外常用的抗Hp药物有羟氨苄青霉素、甲硝唑、克拉霉素、四环素、强力霉素、呋喃唑酮、有机胶态铋剂(De-Nol等)等。溃疡病患者尚可适当结合应用质子泵抑制剂或H2受体拮抗剂加上两种抗菌素,或者质子泵抑制剂(如奥美拉唑)加上一种抗菌素。疗程一般为两个星期。由于治疗Hp感染抗菌方案的广泛应用,有可能扩大耐药性问题的产生。因此,将来替换性的治疗或预防策略,如疫苗预防或免疫治疗的研究是值得重视的。
4.3 Hp感染的流行病学
慢性胃炎患者的胃粘膜活检标本中Hp检出率可达80%~90%,而消化性溃疡患者更高,可达95%以上,甚至接近100%。胃癌由于局部上皮细胞已发生异化,因此检出率高低报道不一。在自然人群中初出生的新生儿血清中抗Hp-IgG水平却很高,接近成人水平,可能从母体获得被动免疫抗体之故。半年后迅速下降。在我国及大多数发展中国家中阳性率待降至10%~20%后又迅速回升。大约在10岁以后即迅速上升达到或接近成人阳性检出率水平。人群Hp感染率因国家因地区有所不同。低达20%,高达90%。人群中总感染率高于发达国家。这些基本资料说明了如下几个问题。①胃病患者中Hp检出率远高于人群中总的检出率,这说明Hp感染者并不都得胃病。这可能还蕴藏着与致病有关的其他因素,特别是遗传因素(宿主的易感性和菌株的型别差异等)。②人群中的Hp感染率与胃病的发生率,发展中国家高于发达国家。这又与社会经济、卫生状况有关。特别是现已证明胃癌高发区不仅与该地区人群中Hp感染率高有关外,还与人群中Hp的早发感染有关。③人类一旦感染Hp后,若不进行治疗,几乎终身处于持续感染中。因此感染率总的讲来随着年龄增长而增长。
关于传染源问题:自然人群中Hp感染率是如此之高,因此是人类Hp感染的主要传染源应该是毫无疑问的。现在的问题是除了人类以外是否还有其他的传染源。在非人的灵长类动物——某些猴类、鼬鼠、猫狗等动物的胃中,亦曾分离到Hp。因此有人认为Hp感染也是动物源性传染病。关于这一点当然不能绝对排除。但是作为传染源来讲这些动物的机会是非常少的,因为人群与这些动物直接接触的机会是不多的(当然属宠物的除外)。
按理Hp是上消化道寄生菌,再加上人群感染率是如此之高,使人们很容易想到其主要传播途径必定是粪—口途径无疑。可是直到如今活的Hp由粪便排出未找到有力的证据。几年前有一位英国学者报告成功地自粪便中分离到Hp,但是其他学者均未重复成功。近年来许多人都是用PCR来证实粪中Hp的存在,结果亦很不一致。有的说粪中阳性率很高,有的说即便在胃中已证实有Hp感染者,粪便中亦未出现阳性。因此粪便中是否有Hp的典型菌,圆球体或残留的HpDNA排出仍是一个谜。可是从有一些人的牙菌斑及龋齿中分离到Hp,为Hp的经口传播提供了一些证据。另外由于消毒不严,由胃镜引起的医源性传播亦已得到证实。上海市医务人员中Hp感染的血清学调查表明从事消化内窥镜工作的医护人员的Hp感染率明显高于其他医务人员。这也可以说是因密切接触后可能造成间接口—口传播的一个证据。
尽管Shahamate等证明Hp能在人工模拟河水的微生态环境中长期存在。并证明在实验室条件下4°C河水中能存活10d。但是迄今未有人从自然环境中分离到Hp。然而Klein等发现胃癌发病率比南美洲高4倍的非洲Lima的普鲁(Peru)族人中,用13CO2呼气试验发现2~12岁儿童中Hp感染者用外源性饮用水(井水)的比用内源性饮用水(自来水)的高3倍。Hulten等进一步对Lima附近一个小镇的饮用水作了研究。他们从Lima取回冰冻水源,用抗Hp-IgG包裹的免疫磁性微球浓缩Hp。用Hp粘附素亚单位基因PCR和Hp约16s rRNa RTPCR(逆转录酶PCR)两种分子生物学手段去检测水源中有无Hp的DNA。结果24/48份标本粘附素亚单位PCR阳性,而11/48份标本两种检测方法均为阳性。因此,他们认为普鲁人饮用水中出现Hp与儿童中流行病学的高发结果是一致的。这为某些条件下可能发生Hp的水源性传播提供了证据。
5 GHLOs与Hp感染的动物模型
5.1 GHLOs
胃螺杆菌样细菌(Gastric Helicobacter likeorganisms GHLOS)是指人胃中发现的Hp以外的在某些性状上类似Hp的细菌,自从1989年Goodwin提出另立Helicobacter一属以来,先后已有许多学者报告了10余种GHLOs,至少包括有雪貂中的H.mustelae(Hm),大鼠和小鼠中的H.muridarum,猫、狗中的H.felis(Hf),卷尾猴中的H.nemetrinae,猎豹中的H.acinonyx,狗中的H.canis和H.bizzozero,小鼠中的H.hepaticus等。另外有人把原属弯曲菌属的有些细菌,亦归入此属。例如H.cinaedi、H.fenneliae和H.heilmanii 。还有人把流产羔羊中分离到的Flexispira rappini,后来在人胃肠类病人中亦发现此菌,且其尿素酶基因与Hp相似,因此亦归入此属,称作H.rappini。这些GHLOs中一部分可能是人或动物胃中的寄生菌,另一部分可能是人或动物胃中的寄生菌,另一部分可能是人或动物胃以外下消化道中的寄生菌。因此它们的生物学性状及致病性等千差万别。除了Hm和Hf以外,目前尚缺少系统的比较资料。
5.2 Hp感染的动物模型
Hp与Hp感染的许多问题的深入解决,尚有待于找到合适的动物模型。但是要建立一个理想的Hp感染动物模型,又是很困难的事。近十年来人们作了很多努力。最早人们用各种常规实验动物作Hp感染均未成功。1987年Krakowka以GF(Germ-free)级小猪用Hp感染,最早建立了悉生(Knotobiotic)动物模型。后来又有人用SPF(Specific-pathogen-frees)级小猎兔犬建立了悉生动物模型,条件非常困难,耗费又大,一般情况下很难办到。因此有人企图用小鼠大鼠等小动物取代之。但直接用Hp感染均未成功,因此人们转而企图用GHLOs(Hf,Hm)在小鼠中建立模拟人类Hp感染的模型。1990年Lee等首先用Hf接种GF级的Swiss Webster小鼠建立了模拟的人类急慢性胃炎的动物模型。后来Karita用BalB/c裸鼠直接感染Hp获得成功。但是裸鼠是一种免疫缺陷型动物,把它用作Hp正常感染情况的研究也有很大缺陷。Fox曾建议用雪貂作动物模型。因为成年的雪貂几乎100%感染了Hm。尽管GHLOs在某些特征上类似于Hp,用它做Hp感染的动物模型能说明一些问题。但GHLOs总终是GHLOs,不能完全代替Hp。裸鼠虽能感染Hp,由于免疫缺陷,亦使其实际应用受到局限。Dubois等又建议用自然感染Hp的族群内繁殖的猴群作Hp感染模型.但这种猴群又往往同时感染有Gastrospillum hominis。总之,十余年来一直未能找到一种十分满意的Hp感染动物模型。最近,Marchetti等用Hp直接感染了CD1和BalB/c,小鼠获得了成功。看来,他们成功的经验在于,Hp原来对这些小鼠的致病力是非常弱的,由于重复通过了这些小鼠,使其致病力有所提高才得以感染。
6 展望
Hp的发现可以说对胃、十二指肠病学的发展产生了一个划时代的影响。近十余年来胃、十二指肠病因学的观念发生了根本的改变,新建立了一系列的侵袭性的和非侵袭性的涉及微生物、病理学、免疫学、生物物理学和分子生物学诊断Hp感染的方法。对急、慢性胃炎及消化性溃疡疾病的治疗改用抗Hp为主的治疗方针后,已取得了良好效果,溃疡的复发率已大大下降。在胃癌与MALT淋巴瘤的发生中也看到了Hp感染在其中起着重要作用,这为今后预防这些肿瘤指明了努力的方向。对Hp本身的生物学性状及其致病机制也已越来越清楚。这些是十余年来取得的巨大进展。然而目前也还存在着一些问题。例如:诊断中最可靠最基础的细菌分离培养工作尚未能普及。准确性较高的非侵袭性13CO2呼气试验及尿15N排出试验等由于涉及设备条件,费用较贵等原因不易推广。快速尿素酶试验及血清学诊断制剂等市场上供应的药盒,种类繁多,缺少统一标准,有的甚至未经过严格鉴定即上市,给临床诊断上造成了混乱。分子生物学方法,亦因种类很多,需要有一定的技术条件,因此还在研究试用阶段,尚未成熟到能普及给临床作出准确诊断的程度。因此,诊断方法的不断改进完善,仍然是不能忽视的一面。慢性胃炎与消化性溃疡由于广泛采用了抗Hp方针为主的治疗后,不可避免地带来越来越严重的毒副作用和耐药性问题。因此,除了不断总结经验,改进治疗方案以外,探讨免疫治疗的可能性及挖掘祖国医学中的伟大宝库亦是一些值得重视的方面。Hp在自然人群中感染的面是如此之广,其涉及的疾病,除了胃癌是我国肿瘤死亡位居第二的病因外,影响广大人民身体健康及劳动生产力的程度也是极其严重的。研究一种像卡介苗一样,能有效地从婴幼儿时期开始即预防这些疾病的疫苗的研究任务已经放在我们面前。希望我们有志者能来突破这一堡垒,为人类作出贡献。当然还有一些问题。例如:Hp感染的传播途径仍需进一步搞清楚。致病机制尚需经合适的动物模型的考核。GHLOs需要好好整理一下,哪些是毫不相干的等等。当前首先一个重要的问题是我们要把Hp的有关知识尽快普及给人民群众,让每家每户都知道:胃病是因传染了Hp后引起的,目前唯一有效的简易办法是讲究卫生,防止“病从口入”,特别是做家长的从婴幼儿出生后即开始注意这一点。若能做到这一点,待有效地疫苗到来之前,我们就已可能从儿童开始减少一大批Hp感染者,大大减少各种胃病的发生率。
张振华
上海第二医科大学微生物学教研室(200025)
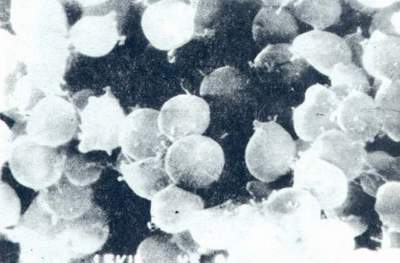
图6 Hp圆球体扫描电镜(×20000)
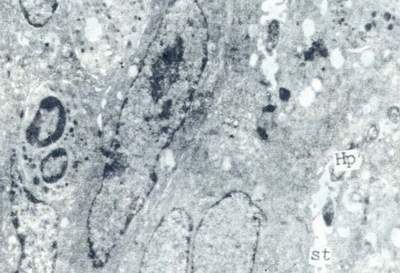
图7 Hp钻入壁细胞的分泌小管中超薄切片,透射电镜(×8000)
Hp:幽门螺杆菌
st:分泌小管
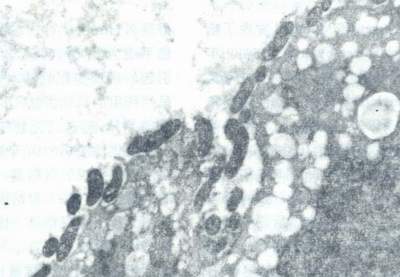
图8 Hp密集地粘附于胃上皮细胞表面超薄切片,透射电镜(×8000)
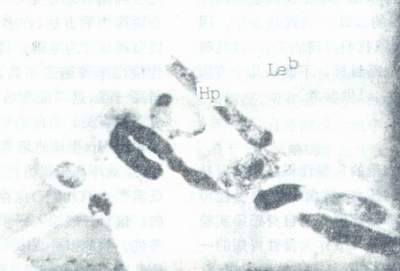
图9 Hp与LebRBC发生的凝集现象超薄切片透射电镜(×12000)
Hp:幽门螺杆菌
Leb:lewisb型红细胞
| 当前页:幽门螺杆菌及其感染 |
| 下一页:我国幽门螺杆菌感染研究的现状和未来 |